Rewriting the Ubiquitin Code: Sugars and Metabolites Enter the Picture
For decades, ubiquitin has been considered almost exclusively as a protein tag. Its C-terminal Gly-Gly motif forms isopeptide (or occasionally thio-/oxy-ester) bonds with Lys, Cys, Ser or Thr residues, marking proteins for degradation, trafficking or regulation. While a handful of reports have suggested that ubiquitin can also be attached to sugars, lipids, nucleic acids or small molecules in vitro, it has been unclear whether such non-protein ubiquitination occurs in cells and in what amounts. One of the main reasons is that standard ubiquitinomic workflows are built around peptides, and carbohydrates or metabolites carrying a ubiquitin chain remain invisible to them.
A new Nature paper from Marco Jochem, Simon A. Cobbold, David Komander and co-workers (WEHI, University of Melbourne, University of Cologne) closes this gap. Their method, Non-Protein Ub-clipping (NoPro-clipping), detects, identifies and quantifies non-proteinaceous ubiquitination directly in cells and tissues, and uncovers a layer of ubiquitin biology that had been inaccessible to systematic study.
A two-enzyme strategy to read out non-protein ubiquitination
NoPro-clipping combines two bacterial enzymes. A Ub-clippase (BpJOS or Lbpro*) cleaves ubiquitin internally, leaving the C-terminal GG dipeptide on the substrate. Sortase A then ligates a biotinylated ClipTag peptide onto the GG remnant, turning the substrate into a peptide-metabolite hybrid amenable to enrichment and LC-MS/MS. For polymeric substrates such as glycogen, samples are pretreated with α-amylase to release short fragments. In vitro standards generated with HOIL-1L (an RBR E3 ligase that forms oxyester linkages on sugars) defined detection limits and diagnostic fragmentation patterns.
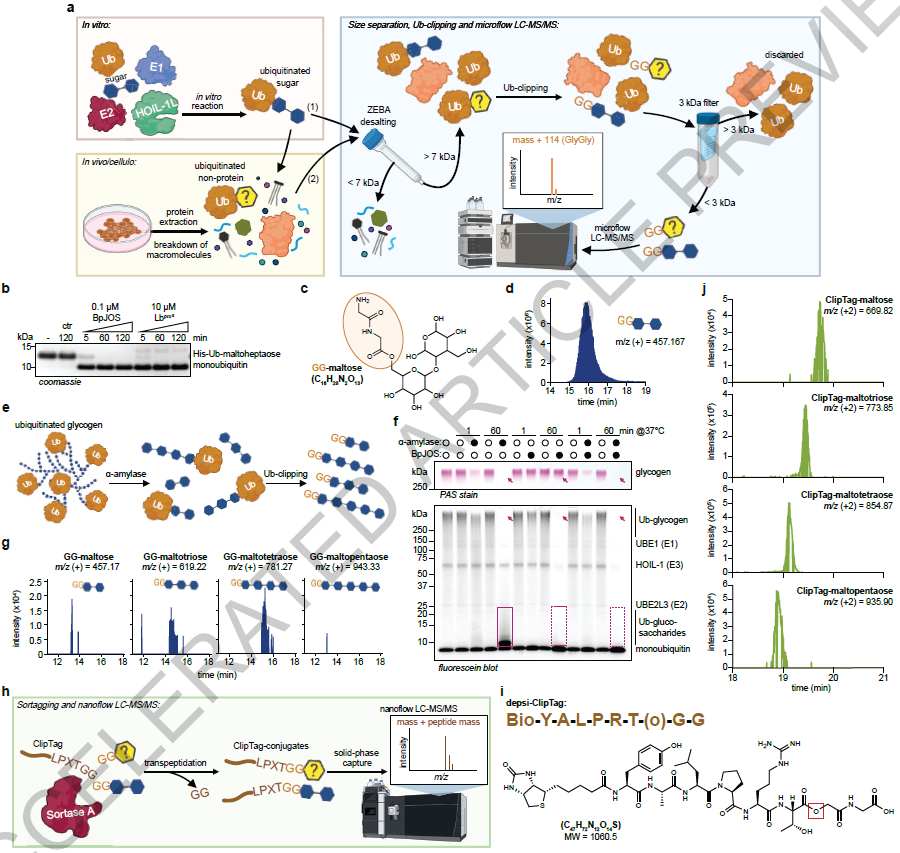
Glycogen ubiquitination by induced proximity
A key section of the manuscript explores the posibility of manipulating glycogen ubiquitination by induced proximity. For that purpose, HOIL-1L and a glycogen-binding domain were fused to complementary dimerisation tags that can be brought together by a small-molecule glue. This allowed the authors to test whether proximity alone drives ubiquitination, see where induced Ub-glycogen ends up in cells, and link ubiquitination to lysosomal trafficking and glycogen clearance. Beyond mechanistic validation, this setup also previews a therapeutic potential, suggesting that induced proximity could be used to drive glycogen ubiquitination and accelerate its clearance.
Ubiquitinated glycogen is widespread and dynamic
Endogenous Ub-glycogen was detected in every glycogen-containing mouse organ examined (liver, skeletal muscle, brain, lung, heart), in human skeletal muscle biopsies, as well as in cancer cell lines.
The most notable observation came from mouse liver. After 6 h of fasting, hepatic glycogen fell by 90% while Ub-glycogen transiently rose, giving an 8-fold increase in the Ub-glycogen to glycogen ratio. Absolute quantification placed Ub-glycogen at over 1% of total cellular ubiquitin, the fourth most abundant ubiquitin conjugate in liver after K48, K63 chains and histone ubiquitination. Refeeding restored glycogen but not Ub-glycogen, indicating that newly synthesised glycogen is initially unmodified. Ubiquitin is therefore a dynamic, nutrition-responsive mark on glycogen.
A route to lysosomal clearance
Lysosomal catabolism of glycogen has been known for over 60 years, but a role for ubiquitin in directing this process had not been established. Ub-glycogen foci colocalised with lysosomes but not endosomes, blocking lysosomal degradation accumulated the signal, and induced ubiquitination reduced detectable cellular glycogen. Ubiquitin therefore acts as a previously missing molecular tag that earmarks glycogen for lysosomal disposal.
A direct link to glycogen storage diseases
The most consequential finding may be the disease connection. In cells modelling GSD IV (loss of the branching enzyme GBE1, producing aberrant polyglucosan aggregates), Ub-glycogen rose 300-fold after normalisation, marking abnormal polyglucosan as a preferred substrate for ubiquitination. Ub-glycogen also depended on the LUBAC complex, which assembles Met1-linked polyubiquitin and whose loss of function causes amylopectinosis (a GSD-like phenotype). Loss of LUBAC activity reduced Ub-glycogen, while loss of OTULIN (the Met1-specific deubiquitinase) raised it. This places Met1-polyubiquitin at the centre of glycogen quality control and provides a long-missing molecular explanation for amylopectinosis.
Beyond glycogen: ubiquitin on small metabolites
Untargeted NoPro-clipping uncovered two further endogenous substrates: glycerol and spermine. Both were validated against in vitro standards, with metabolic labelling and pathway perturbations confirming de novo biosynthesis. Both were also present in mouse liver. Ubiquitin can therefore decorate not only large polymers but also free, central metabolites, hinting at a much broader chemical reach.
My take and limitations
This is a conceptual leap. Ubiquitin is better described as a modifier of biomolecules than as a protein modifier alone, and the link to GSDs and amylopectinosis transforms a curiosity into a candidate disease pathway. If proximity-induced ubiquitination of polyglucosan accelerates lysosomal clearance, this could deliver the first proximity-based therapy for glycogen storage diseases.
A few limitations are worth noting. NoPro-clipping needs catabolic pretreatment tailored to each substrate class, so lipids, nucleotides or cofactors will require dedicated workflows. Species resistant to clipping could be missed, and BpJOS cannot distinguish ubiquitin from NEDD8. The responsible E3 ligases, chain architectures and reader proteins for endogenous Ub-glycogen, Ub-glycerol and Ub-spermine remain unknown, and the latter two could also arise from spontaneous discharging of E1, E2 or E3 thioesters rather than bona fide E3 activity.
Share your opinion
What ubiquitinated biomolecule would you hunt for next with NoPro-clipping? And do you see therapeutic potential in inducing glycogen ubiquitination for GSDs or amylopectinosis?
Full article: https://doi.org/10.1038/s41586-026-10548-x